- +1
演化工程師:生命與環(huán)境如何相互塑造
原創(chuàng) Kevin Laland等 集智俱樂部 收錄于話題#生命復(fù)雜性19個(gè)

導(dǎo)語
在演化的長(zhǎng)河中,生命與環(huán)境持續(xù)地相互塑造。環(huán)境以自然選擇的力量給生物壓力,而生物也會(huì)通過調(diào)整自身所處的生態(tài)位進(jìn)而改變演化壓力,這被稱為生態(tài)位構(gòu)建理論。盡管生態(tài)位構(gòu)建理論尚未成為當(dāng)前主流的演化理論,但其大幅拓展了對(duì)生物適應(yīng)性的理解,并且為人與環(huán)境的關(guān)系、社會(huì)經(jīng)濟(jì)和組織管理等問題提供了新的啟發(fā)。
Kevin Laland、Lynn Chiu | 作者
趙雨亭 | 譯者
張澳 | 審校
鄧一雪 | 編輯
1. 自然選擇:登頂還是移山?
人類正在塑造地球生命演化的未來。我們的活動(dòng)不僅造成大量物種滅絕,還迫使動(dòng)植物與微生物適應(yīng)我們的人造世界——例如,城市鳥類如今改變了自己的鳴叫,因?yàn)楦叩囊粽{(diào)似乎可以使歌聲傳播更遠(yuǎn)。在文化知識(shí)與工程技術(shù)提高人類改造環(huán)境的能力的同時(shí),其它物種也都在改造環(huán)境。在培養(yǎng)皿中涂布細(xì)菌,它們將產(chǎn)生營(yíng)養(yǎng)豐富的代謝產(chǎn)物——新的細(xì)菌菌株可以利用這些副產(chǎn)品,迅速使多種多樣的微生物種群進(jìn)行演化[1]。海貍創(chuàng)造的池塘為鴨子提供了繁殖地;殘留的蜘蛛網(wǎng)被地上的昆蟲開發(fā)為冬眠地;植物通過其根部分泌的物質(zhì)來改造土壤。這些生物改變環(huán)境的方式反過來改變了它們和其他物種在生存與繁殖中所面臨的演化壓力。換句話說,它們的行為使自然選擇產(chǎn)生偏向。所有物種都會(huì)進(jìn)行這種被稱為“生態(tài)位構(gòu)建(niche construction)”的過程,雖然它們?cè)斐傻挠绊戇h(yuǎn)不如人類劇烈而廣泛。
然而,生態(tài)位構(gòu)建的理論[2]在演化生物學(xué)家中引起爭(zhēng)議,部分原因是傳統(tǒng)上認(rèn)為自然選擇是“盲目的”:人們認(rèn)為,對(duì)生物體上千年的調(diào)整是為了使其適應(yīng)各自的生態(tài)位,而不是為了達(dá)成生物體各自的目標(biāo)。人類經(jīng)歷了同樣的改造,但相比于為了適應(yīng)預(yù)設(shè)的生態(tài)位而演化,我們更加主動(dòng),更像是在改造環(huán)境而使其適于生存。我們的大腦已經(jīng)演化為適合處理語言編碼的信息與知識(shí),因?yàn)槲覀兘⒘素S富的文化領(lǐng)域,并隨后對(duì)其適應(yīng)。我們馴化了動(dòng)植物,將其納入我們的食物中,從而觸發(fā)了代謝此類食物的基因選擇。我們創(chuàng)造了農(nóng)業(yè),促使人口增長(zhǎng),無意中篩選了對(duì)群體疾病(如傷寒或霍亂)的抵抗物質(zhì)。我們的父輩不僅將基因傳遞給我們,同時(shí)還留下了其改造后的世界。這種生態(tài)遺產(chǎn)意味著人類并非響應(yīng)靜態(tài)的環(huán)境而演化,而是塑造該環(huán)境以緩解或加劇其施加的某種選擇壓力。演化生物學(xué)家理查德·萊文斯(Richard Levins)和理查德·列旺汀(Richard Lewontin)指出[3],“有機(jī)體既是自然選擇的對(duì)象,又是選擇條件的創(chuàng)造者,從而影響著自身的演化。”
但這種觀點(diǎn)仍有爭(zhēng)議:專家們對(duì)其它生物通過改變環(huán)境來指導(dǎo)演化的程度產(chǎn)生了分歧。地形的比喻可以幫助理解演化過程。我們可以將演化視為適應(yīng)性景觀(adaptive landscapes,也譯為適應(yīng)性地形圖),該圖揭示了生物體的適應(yīng)性(其生存和繁殖能力)與一種或多種生物體性狀之間的關(guān)系。例如,在一張鳥群適應(yīng)性景觀中,一個(gè)軸可能代表體型大小,另一個(gè)軸可能代表尾巴長(zhǎng)度(或者可以用相關(guān)基因的頻率來替代)。該地形圖為三維空間中的曲面,具有更高適應(yīng)性的鳥——即得分最高的那些——具有更優(yōu)良的體型與尾巴長(zhǎng)度組合,而適應(yīng)性較差的鳥則處于曲線低谷。曲面最高峰的區(qū)域代表具有最佳體型和尾巴長(zhǎng)度的鳥,同時(shí)也是生存能力最強(qiáng)的鳥。

圖1. 三維的適應(yīng)性景觀
從演化尺度上看,鳥類種群具有一系列特征,每種組合在地形圖中對(duì)應(yīng)不同的點(diǎn)——通過自然選擇,種群將逐漸收斂到最適合當(dāng)?shù)貤⒌氐奶卣鳎ㄟ^種群演化而在適應(yīng)性景觀上攀升至適應(yīng)性的局部高峰。然而登頂之路不止一條:可以調(diào)整自己的性狀以到達(dá)頂峰,也可以移動(dòng)山峰以適應(yīng)自身的需求。

圖2. 一種不受生物影響的靜態(tài)地形圖。在圖示中,時(shí)間向下移動(dòng)。改自 Tanaka et al (2020), Philosophy of Science, 87: 478-98
2. 蚯蚓的進(jìn)擊:從適應(yīng)到入侵
小巧而有力的生態(tài)位構(gòu)造者隨處可見。低調(diào)的蚯蚓就是一位“生態(tài)系統(tǒng)工程師[4]”,它是一種會(huì)改變物理與化學(xué)環(huán)境的物種,并常以改變生態(tài)系統(tǒng)功能的方式改造環(huán)境。奇怪的是,盡管在生態(tài)學(xué)上蚯蚓是成功的,但在解剖學(xué)上,它們實(shí)際并不適合在陸地上生存。盡管已經(jīng)在陸地上生活了數(shù)千萬年(甚至可能數(shù)億年),但它們?nèi)员A糁鴱牡醒莼鴣淼幕旧硖匦浴Ec大多數(shù)陸生動(dòng)物相異,蚯蚓會(huì)如淡水動(dòng)物般代謝產(chǎn)生大量尿液,這使它們極易脫水而亡——如果在晴天挖出了蚯蚓,它們通常會(huì)在短時(shí)間內(nèi)干枯。那么,這樣一種生理適應(yīng)性如此差的動(dòng)物是如何在陸地上生存這么長(zhǎng)時(shí)間的呢?
答案是生態(tài)位構(gòu)建——相比于調(diào)整生理特征以適應(yīng)陸生環(huán)境,它們選擇了在陸地上制造水生世界。它們的成功直接源于對(duì)自然所干的園藝活。蚯蚓通過松土減少土壤的“基質(zhì)潛力”——即土壤保持水的能力。這使蚯蚓更容易通過周圍環(huán)境攝取水分,以補(bǔ)充由于它們的排泄而造成的水分損失。換句話說,蚯蚓有效地建立了自己的儲(chǔ)水池[5],使自己保持濕潤(rùn)。
就這樣,蚯蚓發(fā)展了許多將環(huán)境改造為適應(yīng)其自身發(fā)展的方式,并造成了與人類入侵自然類似的影響。如今歐洲蚯蚓被認(rèn)為是北美森林植物群落發(fā)生變化的主要推動(dòng)力[6]——并且與人們認(rèn)為蚯蚓對(duì)土壤有益的固有印象相反,蚯蚓對(duì)闊葉林具有毀滅性的影響。北美本土蚯蚓種群在第四紀(jì)冰期期間幾乎滅絕,在16世紀(jì)和17世紀(jì)歐洲殖民者將蚯蚓引入前,多數(shù)生態(tài)系統(tǒng)沒有蚯蚓的存在。隨后,歐洲蚯蚓迅速擴(kuò)散,它們?nèi)肭致淙~林并快速改變了土壤性質(zhì),使本地動(dòng)植物物種生存困難。
3. 入侵導(dǎo)致的級(jí)聯(lián)反應(yīng)
蚯蚓將有機(jī)物作為食物或用作挖洞的材料拖入土壤中,導(dǎo)致枯枝落葉層迅速流失,并導(dǎo)致土壤表面升溫、脫水。蚯蚓的活動(dòng)改變了碳和氮等化學(xué)元素在整個(gè)生態(tài)系統(tǒng)中循環(huán)的速率——例如,加快了植物有機(jī)體的分解速度,同時(shí)它們的挖掘行為提升了土壤的通氣和排水效果。這些變化通常在入侵的前十年內(nèi)發(fā)生,進(jìn)而引發(fā)了持續(xù)數(shù)十年的演替狀態(tài)。歷史上,在那些沒有蚯蚓的森林中,大量碳元素被鎖于森林表面的枯枝落葉層下。而當(dāng)蚯蚓消耗這些枯枝落葉時(shí),大量?jī)?chǔ)存的碳元素以二氧化碳的形式被釋放到大氣中,就像人類使用化石燃料一樣,從而導(dǎo)致全球變暖[7]。

圖3. 被稱為“森林地板”的枯枝落葉層,其中包含大量的有機(jī)物質(zhì)
遭遇蚯蚓入侵的棲息地產(chǎn)生了方方面面的變化,對(duì)其它物種產(chǎn)生了深遠(yuǎn)的影響。最近的研究發(fā)現(xiàn)[8],隨著引入的蚯蚓種群豐富性增加,植物多樣性下降,其中本土植物受到的沖擊最大;隨著蚯蚓生物量的增加,土壤無脊椎動(dòng)物減少[9]。本土動(dòng)植物遭受沖擊的原因是因?yàn)樗鼈円酝m應(yīng)的是不存在蚯蚓的環(huán)境。同時(shí),某些物種受益:就像人類的侵?jǐn)_一樣,蚯蚓的工程促進(jìn)了快速生長(zhǎng)的細(xì)菌、草本植物與一些大型動(dòng)物(例如鹿)的入侵和繁殖。蚯蚓的洞穴被其它無脊椎動(dòng)物——甚至某些脊椎動(dòng)物(如蠑螈)——用作巢穴[10],而這些生物的糞便為微生物活動(dòng)的提供了充足條件。與蚯蚓同時(shí)出現(xiàn)的往往還有侵入性灌木,如沙棘和金銀花。部分原因是蚯蚓通過選擇性地(例如根據(jù)種子大小等性狀)食用種子,改變了種子庫(kù)(種子在土壤中的自然儲(chǔ)存)的組成。從蚯蚓口中存活下來的種子隨后受益于蚯蚓糞便提供的營(yíng)養(yǎng)及物理保護(hù),獲益于此的依然是非本土植物。
蚯蚓的入侵無疑會(huì)觸發(fā)森林生態(tài)系統(tǒng)的級(jí)聯(lián)反應(yīng)。一些研究人員甚至擔(dān)心蚯蚓可能引發(fā)“入侵崩潰(invasion meltdown)[11]”,其中非本土物種加速促進(jìn)彼此的入侵[12]。盡管不夠直觀,但蚯蚓自身會(huì)從其工程設(shè)計(jì)及其觸發(fā)的入侵中受益,從而使它們能夠在適應(yīng)性景觀上創(chuàng)造新的山峰。蚯蚓促進(jìn)侵入性灌木的生長(zhǎng),侵入性灌木反過來促進(jìn)蚯蚓種群的生長(zhǎng)[13]。灌木產(chǎn)生更高質(zhì)的枯枝落葉層(即較低的碳氮比)以供歐洲蚯蚓享用,使自持續(xù)的反饋閉環(huán)得以形成。蚯蚓積極參與耕作——準(zhǔn)備土壤,改變種子庫(kù),使得其偏愛的作物肆意生長(zhǎng)。它們與鹿、豬和微生物建立了類似的互惠關(guān)系,后者既從蚯蚓的活動(dòng)中受益,也促進(jìn)了蚯蚓的活動(dòng)。有趣的是,蚯蚓物種的多樣性與其生物量相關(guān)[14],這表明某一種蚯蚓的工程對(duì)其它各種蚯蚓均有益。
4. 生態(tài)遺產(chǎn)的累積
蚯蚓也會(huì)通過建造洞穴來積累演化上上的優(yōu)勢(shì)。目前地球上有6000多種蚯蚓,它們?cè)谏鷳B(tài)和行為上有很大不同。然而,大多數(shù)蚯蚓并不是閑逛的“游民”,在土地上四處游蕩以尋找食物, 而將遺棄的土地扔在身后。相反,蚯蚓可以建造半永久的洞穴,其存留時(shí)間通常遠(yuǎn)超蚯蚓的一生。查爾斯·達(dá)爾文曾指出,蚯蚓的洞穴并不是簡(jiǎn)單的挖掘工作,而是堅(jiān)固的垂直隧道,其深度有時(shí)超過一米,他將其描述為“內(nèi)襯水泥”。蚯蚓的“水泥”通過壓實(shí)其分泌物形成[15],并創(chuàng)造了厚度足有一厘米的獨(dú)特洞穴,稱為“蚓觸圈(drilosphere)”。每一個(gè)蚯蚓的洞穴(它們的生活區(qū))會(huì)在土壤表面形成糞堆,既是廚房又是廁所,由枯枝落葉及其排泄物組成。蚯蚓甚至?xí)诰蛴糜诋a(chǎn)卵的側(cè)洞。許多蚯蚓在夜間尋找食物和配偶,它們通常把尾巴末端置于洞內(nèi),以便快速響應(yīng)危機(jī)。

圖4. 蚯蚓錯(cuò)綜復(fù)雜的洞穴
挖洞是一項(xiàng)耗費(fèi)精力的活動(dòng)——認(rèn)領(lǐng)和翻修前任主人的房子要比從頭開始建造自己的房子容易得多。人們通常認(rèn)為蚯蚓挖的洞穴比它們自身寬度窄,但常能在大洞穴里發(fā)現(xiàn)小蚯蚓,這表明要么是蚯蚓幼蟲繼承了父母的洞穴,要么是分散的個(gè)體傾向于在空的洞穴里安家。長(zhǎng)期的野外觀察表明,洞穴被使用的時(shí)間遠(yuǎn)超蚯蚓一生的生命。因此,蚯蚓在被使用過的土壤與未被開墾的土壤中更傾向于前者也不足為奇了[16]。
改良的土壤、現(xiàn)成的房屋和更充分的食物供應(yīng),并不是蚯蚓留給后代的全部。每一個(gè)蚯蚓繭都繼承了一個(gè)來自其父母腸道的微生物群落。蚯蚓的環(huán)境不僅僅是土壤和周圍的生物,還有寄居在其消化系統(tǒng)和周圍蚓觸圈中的細(xì)菌、真菌和單細(xì)胞真核生物。這種安排同樣是互利的:蚯蚓為土壤微生物提供了一個(gè)食物豐富和可移動(dòng)的環(huán)境,讓它們有機(jī)會(huì)在缺氧的穴中發(fā)酵并在土壤中繁殖,而這些共生體的存在增加了蚯蚓的適應(yīng)性[17]。
就像人類一樣,蚯蚓已經(jīng)建立了自己的生態(tài)位。它們的生活環(huán)境并不是一直保持貧瘠或富庶,而是根據(jù)其內(nèi)棲息者的活動(dòng)呈現(xiàn)動(dòng)態(tài)的變化。除了改變土壤,蚯蚓還如人類一般種植農(nóng)作物、釋放溫室氣體、形成聚落并向外拓展。
5. 生態(tài)位構(gòu)建:刻意的設(shè)計(jì)
蚯蚓并不是通過生態(tài)位構(gòu)建改善環(huán)境的唯一物種:海藻分泌的粘性化學(xué)物質(zhì)會(huì)束縛沙土并穩(wěn)定環(huán)境[18];叢林和松樹通過散布易燃的針葉、果皮和油類物質(zhì)來提升森林火災(zāi)規(guī)模[19],從而消除競(jìng)爭(zhēng);螞蟻則通過噴灑酸性除草劑或種植真菌作物[20],消除不適合其居住生活的樹木[21]。
動(dòng)物根據(jù)自己的需求改善環(huán)境的方式是否激勵(lì)了我們以新的方式思考演化的適應(yīng)性景觀?從最根本的生態(tài)位構(gòu)建的角度來看[22],答案是“YES”:生物不單單適應(yīng)環(huán)境,生物與環(huán)境兩者共同指導(dǎo)演化。例如,海貍的影響要比它們創(chuàng)造的池塘要廣泛得多。海貍既是消防員[23],同時(shí)也是灌溉管理人員[24]——它們極大地改變了所在的生態(tài)系統(tǒng),使其能夠抵御諸如火災(zāi)或是干旱等惡劣氣候的干擾。這樣,海貍便會(huì)改變其本地環(huán)境,從而改變選擇對(duì)它們和其他物種的作用方式。海貍是反饋與因果關(guān)系循環(huán)網(wǎng)絡(luò)的一部分,在該網(wǎng)絡(luò)中,初期占據(jù)優(yōu)勢(shì)的生物體會(huì)驅(qū)動(dòng)環(huán)境變化,而經(jīng)優(yōu)勢(shì)物種調(diào)整的環(huán)境隨后會(huì)選擇生物體的演化方向。
生態(tài)位構(gòu)建與生態(tài)和演化中的其他幾個(gè)概念重疊,特別是“生態(tài)系統(tǒng)工程”和“生態(tài)演化動(dòng)力學(xué)”。但是,生態(tài)位構(gòu)建理論的獨(dú)特之處在于,生物對(duì)環(huán)境的改造(生態(tài)位構(gòu)建)及其對(duì)后世的遺產(chǎn)(生態(tài)繼承)是演化過程,正是它們引發(fā)了演化的改變。
盡管大多數(shù)教科書都側(cè)重于描述直接改變基因頻率的現(xiàn)象——例如自然選擇、突變或隨機(jī)遺傳漂變(偶然發(fā)生的群體遺傳組成變化),演化過程的起因目前仍有爭(zhēng)議。回到適應(yīng)性景觀的比喻,這些力量推動(dòng)著生物群在環(huán)境中的適應(yīng)性上下移動(dòng)。相反,生態(tài)位構(gòu)建理論則提倡更廣泛的演化因果關(guān)系概念。
引用演化生物學(xué)家道格拉斯·富圖瑪(Douglas Futuyma)的話說[25],“傳統(tǒng)主義者”會(huì)說,盡管生態(tài)位構(gòu)建“可以影響甚至導(dǎo)致自然選擇的演化過程”;但“生態(tài)位構(gòu)建自身并非演化過程,最多就是一種變化的環(huán)境”。這種觀點(diǎn)有所欠缺:更一般地將生態(tài)位構(gòu)建與環(huán)境變化區(qū)分開的是,生態(tài)位構(gòu)建以有序、有針對(duì)性與持續(xù)的方式改變了自然選擇。生態(tài)位構(gòu)建不僅僅會(huì)改變自然選擇,而且還會(huì)通過對(duì)選擇的方向和強(qiáng)度施加統(tǒng)計(jì)偏差來指導(dǎo)自適應(yīng)演化。生物對(duì)適應(yīng)性景觀進(jìn)行設(shè)計(jì)。例如,對(duì)美國(guó)東北部15個(gè)森林中原生植物群落上生物入侵進(jìn)行的分析顯示出驚人的一致模式。盡管蚯蚓種類和豐度、本地植物組成與特性、以及地理區(qū)域存在差異,但在所有地點(diǎn)都檢測(cè)到了與蚯蚓相關(guān)的相同反應(yīng)。
如Lewontin所建議的那樣,相比于將演化視為適應(yīng)性景觀,將其視為生物走過蹦床的過程可能會(huì)更好。但是,已有示例顯示,隨著生物在演化過程中的變遷,它們不僅會(huì)抑制自身的適應(yīng)能力,而且還可以改善環(huán)境,從而改善其適應(yīng)能力。該過程強(qiáng)大到可以影響演化結(jié)果的確定。例如,蚯蚓并沒有上升到典型的陸生動(dòng)物生理的適應(yīng)高峰。它們反而又進(jìn)行了一次演化旅程,在陸地到達(dá)了一個(gè)難以置信的水生動(dòng)物生理適應(yīng)高峰。

圖5. 被生物抑制的適應(yīng)性景觀。在圖示中,時(shí)間向下移動(dòng)。圖改自Tanaka et al (2020), Philosophy of Science, 87: 478-98。
6. 相互作用的地形圖
最近,Mark Tanaka、Peter Godfrey-Smith和Benjamin Kerr——一個(gè)理論生物學(xué)家與哲學(xué)家組合的團(tuán)隊(duì)——設(shè)計(jì)了一種新穎的數(shù)學(xué)框架[26],用于對(duì)自然選擇和生態(tài)位構(gòu)建進(jìn)行建模。他們以兩個(gè)關(guān)聯(lián)的地形共同描述適應(yīng)性演化,而不是僅用單一的適應(yīng)性曲面:有機(jī)體通過在適應(yīng)性(adaptive)景觀上攀登而演化,同時(shí)環(huán)境的演化是通過在構(gòu)建性(constructive)景觀上攀登而發(fā)生的。兩者之間的關(guān)系決定了特定生物的成功程度。就好像不追蹤雌孔雀擇偶偏好的演變,就無法充分了解雄孔雀尾巴這類第二性征的演化。因此,如果不考慮生態(tài)位的構(gòu)建,就無法描述適應(yīng)性隨生物對(duì)環(huán)境特征的改變所產(chǎn)生的演變。Tanaka和同事使用他們的生態(tài)位構(gòu)建演化框架來分析三種微生物演化案例的動(dòng)力學(xué),其中微生物參與了不同形式的生態(tài)位構(gòu)建。
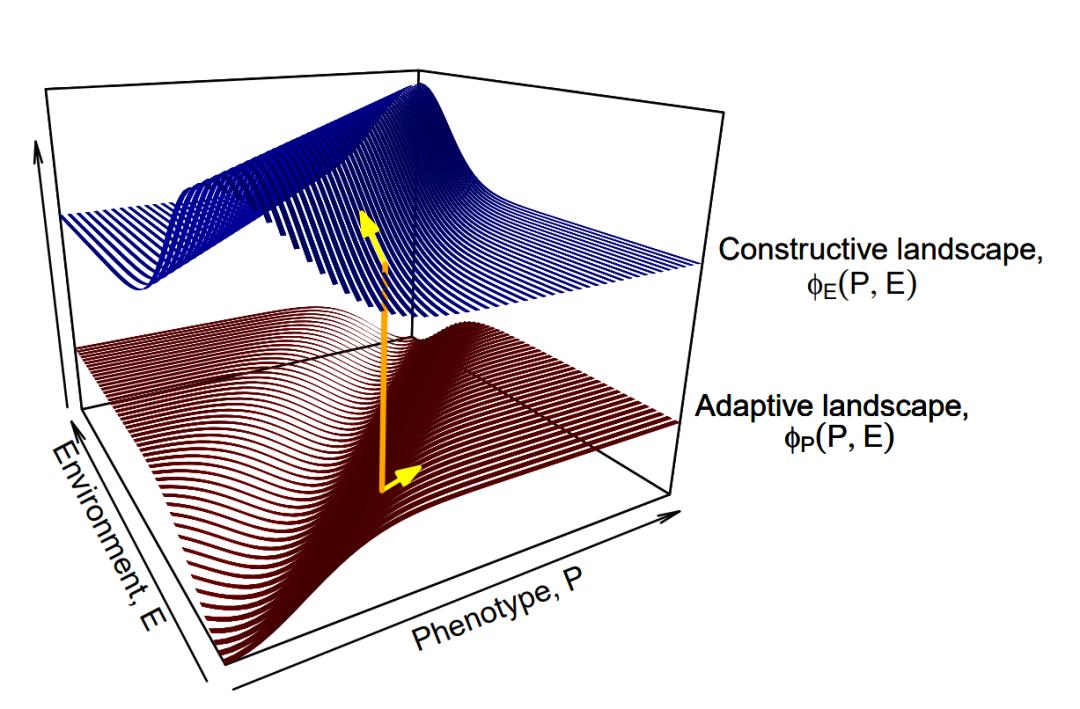
圖6. 適應(yīng)性景觀(下部紅色曲面)與建設(shè)性地形圖(上部藍(lán)色曲面)。黃色的向量箭頭指示了在(P,E)點(diǎn)上表現(xiàn)型(Phenotype)方向(下圖)與環(huán)境(Environment)方向(上圖)上的偏微分。橙色的豎線將兩向量相連。
所謂的費(fèi)舍爾基本定理(Fisher's fundamental theorem)描述了自然選擇推動(dòng)種群向改變自身來適應(yīng)地形的方式:通過自然選擇演化的種群適應(yīng)性的增長(zhǎng)率等于適應(yīng)性的遺傳方差(即導(dǎo)致生存及繁衍能力產(chǎn)生變化的遺傳變異量),從而導(dǎo)致生存和繁殖差異。統(tǒng)計(jì)學(xué)家兼遺傳學(xué)家羅納德·費(fèi)舍爾(Ronald Fisher)認(rèn)為,由于種群適應(yīng)性無法無限增長(zhǎng),因此必須考慮環(huán)境的惡化來平衡這一過程。Tanaka和他的同事們構(gòu)建的地形耦合模型從數(shù)學(xué)上表明,在生態(tài)位構(gòu)建中,這種適應(yīng)性的守恒不一定存在。生物可以通過改變其環(huán)境的方式來提高其絕對(duì)適應(yīng)性(如在山峰筑丘),同時(shí)也可以破壞其環(huán)境并降低其適應(yīng)性(如挖坑)。
該模型和解釋了蚯蚓與其他重塑環(huán)境的物種(包括人類)的演化有關(guān)。歐洲蚯蚓在北美森林中取得的巨大成功,很可能不是自然選擇使其適應(yīng)森林環(huán)境的表現(xiàn),而是它們改變森林生態(tài)以使森林更適應(yīng)自身能力的結(jié)果。對(duì)于人類而言,這一點(diǎn)也適用。馴化植物、耕種土壤、采用新的灌溉方法和施用肥料——它們都增加了土地供養(yǎng)人類的能力。通過反復(fù)改善環(huán)境,我們已經(jīng)提高了我們的絕對(duì)適應(yīng)能力,這就好似我們?cè)谶m應(yīng)性景觀上建造了越來越高的塔,然后爬上塔來到達(dá)地形的巔峰。
7. 性狀演化:自然選擇+生態(tài)遺產(chǎn)
演化生態(tài)學(xué)家Tobias Uller 與 Heikki Helanter?設(shè)計(jì)了一種方法,將生態(tài)位構(gòu)建結(jié)合到另一個(gè)普遍的演化模型中,即以群體遺傳學(xué)家喬治·普萊斯(George Price)命名的普萊斯方程[27]。該模型建立在兩個(gè)核心的演化過程之上:對(duì)性狀的自然選擇隨時(shí)間的演化;以及性狀在幾代之間有偏好性的傳遞(或繼承)。這些過程中的每一步都可以進(jìn)行統(tǒng)計(jì)定義和測(cè)量,從而提供一種計(jì)算性狀隨時(shí)間變化(即演化)以及選擇或傳遞貢獻(xiàn)量的方法。自然選擇項(xiàng)由性狀的可遺傳性(遺傳差異導(dǎo)致性狀差異的程度)與選擇差異(衡量性狀對(duì)適應(yīng)性的影響程度)的乘積給出。例如,如果大型狗相比小型狗擁有更多的幼狗,但是耳朵直立的狗與垂耳的狗后代數(shù)量相似,那么體型大小會(huì)具有較大的選擇差異,而耳朵形狀的選擇差異將會(huì)很小。感興趣的性狀可以是任何事物,包括對(duì)環(huán)境的適應(yīng)程度。由于遺傳力與遺傳變異的數(shù)量有關(guān),因此當(dāng)關(guān)注的性狀是適應(yīng)性時(shí),普萊斯方程中的自然選擇項(xiàng)廣義上與費(fèi)舍爾基本定理等價(jià)。

圖7. 普萊斯方程模型模擬了自然選擇(可以分解為兩個(gè)組成部分:遺傳力和選擇差異)和非選擇因素(如傳遞偏差)導(dǎo)致的演化變化。生態(tài)位的建設(shè)可以影響演化的所有三個(gè)組成部分,蚯蚓就是一個(gè)很好的例子。圖片由作者提供
相比之下,性狀傳遞項(xiàng)代表的是自然選擇之外的其他因素,這些因素會(huì)影響性狀隨時(shí)間的變化,包括其對(duì)遺傳何種性狀的影響。有趣的是,Price將其稱為“環(huán)境變化項(xiàng)”,這似乎特別適合于此類場(chǎng)景。蚯蚓并不是隨意地改變土壤——有時(shí)會(huì)給后代留下一個(gè)更好的世界,有時(shí)會(huì)變得更糟,它們也沒有持續(xù)分解土壤。相反,蚯蚓一代又一代地提高了土壤的質(zhì)量,通過生態(tài)位的構(gòu)建提高了自身的適應(yīng)能力,并為其后代留下了有偏好性的生態(tài)遺產(chǎn)。正如費(fèi)舍爾(Fisher)認(rèn)為環(huán)境應(yīng)該惡化的假設(shè)一樣,歷史上,當(dāng)關(guān)注的性狀為適應(yīng)性時(shí),研究人員傾向于認(rèn)為普萊斯方程中傳遞偏差項(xiàng)為負(fù)值。這是因?yàn)橥蛔兺ǔ?duì)生物不利,也因?yàn)榄h(huán)境影響被(錯(cuò)誤地)認(rèn)為不會(huì)被遺傳。結(jié)果,與傳遞項(xiàng)有關(guān)的過程在演化中所起的作用相當(dāng)程度上已被忽略。當(dāng)有機(jī)體為其后代構(gòu)建或選擇更好的環(huán)境時(shí),即使沒有自然選擇,它們也可能引起適應(yīng)性演化[28]。
Uller和Helanter?使用普萊斯方程指出了生態(tài)位構(gòu)建可以影響演化的其他方式[29]。在演化分析中,可遺傳性(由于遺傳差異而導(dǎo)致的性狀差異的多少)通常是通過統(tǒng)計(jì)(例如,父輩與子輩間的回歸)來估計(jì)的,該統(tǒng)計(jì)數(shù)據(jù)比較了父母和后代的性狀相似性(例如,大狗的幼犬是否更大?)。父輩將改造后的環(huán)境傳遞給后代,使得可遺傳性的估計(jì)值偏大。
對(duì)該可遺傳性估值偏大的現(xiàn)象,蚯蚓又一次提供了有用的例證——這次是是粘液——因?yàn)樗鼈兎置谡骋阂员Wo(hù)其皮膚外層,并且粘液的分泌對(duì)土壤特性十分敏感[30]。在蚯蚓處理程度不同的土壤條件下進(jìn)行測(cè)量,蚯蚓父輩與子輩之間的粘液水平之間存在很強(qiáng)的相關(guān)性。但是,這并不是因?yàn)轵球緯?huì)將“多余的粘液”基因傳給嬰兒。親子后代的相似性和高遺傳力的出現(xiàn)主要是由于生態(tài)學(xué)而不是遺傳遺傳:后代“繼承”了其父母的土壤(物質(zhì)環(huán)境與微生物組),因此產(chǎn)生了類似數(shù)量的粘液。如果研究人員調(diào)查粘液水平如何對(duì)入侵的蚯蚓進(jìn)行自然選擇,那么他們很可能會(huì)檢測(cè)到性狀發(fā)生強(qiáng)烈變化,這意味著選擇低量的粘液分泌。但實(shí)際上,這是生態(tài)位構(gòu)建的生態(tài)效應(yīng),因?yàn)轵球居H本已經(jīng)為后代改善了土壤環(huán)境。目前尚無對(duì)自然選擇的遺傳反應(yīng),但性狀發(fā)生了系統(tǒng)性和方向性變化,表明性狀與環(huán)境適應(yīng)性密切相關(guān)。
8. 適應(yīng)性蹺蹺板
生態(tài)位構(gòu)建對(duì)演化的另一種影響方式是改變性狀影響適應(yīng)性的方式(即改變選擇差異,這是普萊斯方程中自然選擇項(xiàng)的一部分)。例如,對(duì)于蚯蚓遇到貧瘠土壤及食物缺乏傾向于遷徙的性狀[31],實(shí)驗(yàn)研究表明,可以通過改善棲息地質(zhì)量來減少自身(和其他蚯蚓物種)遷徙的需求[32]。它們沒有遷移到更富裕的環(huán)境中——而是灌溉、施肥和重新種植土地,將其轉(zhuǎn)變?yōu)閮?yōu)質(zhì)土壤。
現(xiàn)在考慮在入侵的蚯蚓種群中,存在“快速”和“慢速”兩種擴(kuò)散者。擁有“快速擴(kuò)散”基因的蚯蚓可能會(huì)定居在土壤中,但由于原始土壤的質(zhì)量較差,因此最初適應(yīng)性較低。但隨著它們努力改善環(huán)境,更多的無脊椎物種以及有益的侵入性灌木和動(dòng)物(對(duì)它們有益)被吸引,因此它們的適應(yīng)性進(jìn)一步提高。最終,蚯蚓種群變得密集,以至于其適應(yīng)性因競(jìng)爭(zhēng)和資源過度開發(fā)而下降,從而引發(fā)了新一輪的傳播浪潮。相比之下,擁有“慢速擴(kuò)散”基因的蚯蚓最初比定居者具有更高的相對(duì)適應(yīng)性。但是,隨著被入侵森林土壤質(zhì)量的改善,慢速擴(kuò)散的蚯蚓相對(duì)適應(yīng)性會(huì)下降,直到它們過度分散為止。在生態(tài)位構(gòu)建和競(jìng)爭(zhēng)的推動(dòng)下,快速和緩慢的擴(kuò)散者陷入了周期性的戰(zhàn)斗。回到適應(yīng)性景觀的比喻,這里的地形就像一個(gè)蹺蹺板:慢速擴(kuò)散受到青睞,但隨著快速擴(kuò)散者改善土壤,蹺蹺板向另一端傾斜,而快速擴(kuò)散則成為適應(yīng)性時(shí)的最高點(diǎn),直到蹺蹺板再次翻轉(zhuǎn)。

圖8. 適應(yīng)性蹺蹺板,適應(yīng)的方向因生物而改變。在圖示中,時(shí)間向下移動(dòng)。圖改自Tanaka et al (2020), Philosophy of Science, 87: 478-98
所有這些形式化的數(shù)學(xué)分析都有助于加深對(duì)生物改變環(huán)境后果的理解。為了理解適應(yīng)性演化,研究人員不僅需要了解生態(tài)位構(gòu)建如何通過自然選擇而演化,還需要了解生態(tài)位構(gòu)建如何改變自然選擇的環(huán)境來源。我們需要擴(kuò)展適應(yīng)性景觀的比喻,以刻畫由生物所引導(dǎo)的環(huán)境變化。不斷演化的種群不像僵尸登山者那樣盲目地攀登適應(yīng)性高峰,而是更像勤勞的地形設(shè)計(jì)師。它們配備了挖掘和建造設(shè)備,可以根據(jù)自己的需求對(duì)地形進(jìn)行重塑。在人類生態(tài)位構(gòu)建和生態(tài)遺產(chǎn)正在破壞地球的生態(tài)并推動(dòng)人口爆炸增長(zhǎng)的現(xiàn)狀下,了解生物體如何通過重構(gòu)生態(tài)而提高適應(yīng)性變得更加緊迫。
原文鏈接:
https://aeon.co/essays/organisms-are-not-passive-recipients-of-evolutionary-forces
參考資料
[1]https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1006340
[2]https://nicheconstruction.com/
[3] https://www.hup.harvard.edu/catalog.php?isbn=9780674202832
[4]https://www.hup.harvard.edu/catalog.php?isbn=9780674202832
[5]https://www.esf.edu/efb/turner/pubsPages/extendedorganism.html
[6]https://conbio.onlinelibrary.wiley.com/doi/abs/10.1111/j.1523-1739.2009.01168.x
[7]https://www.nature.com/articles/nclimate1692
[8]https://onlinelibrary.wiley.com/doi/10.1111/gcb.13446
[9]https://besjournals.onlinelibrary.wiley.com/doi/full/10.1111/1365-2656.12746
[10]https://link.springer.com/article/10.1007/s10530-006-9021-9
[11]https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6944502/
[12]https://experts.umn.edu/en/publications/european-buckthorn-and-asian-soybean-aphid-as-components-of-an-ex
[13]https://link.springer.com/article/10.1007/s10530-008-9281-7
[14]https://onlinelibrary.wiley.com/doi/10.1111/gcb.13446
[15]https://link.springer.com/chapter/10.1007%2F978-3-642-14636-7_8
[16]https://link.springer.com/chapter/10.1007%2F978-3-642-14636-7_80
[17]https://link.springer.com/article/10.1007%2Fs00374-009-0393-8
[18]https://pubmed.ncbi.nlm.nih.gov/20511426/
[19]https://academic.oup.com/bioscience/article/56/7/570/234327
[20]https://www.journals.uchicago.edu/doi/10.1086/379351
[21]https://www.nature.com/articles/437495a
[22]https://press.princeton.edu/books/paperback/9780691044378/niche-construction
[23]https://esajournals.onlinelibrary.wiley.com/doi/epdf/10.1002/eap.2225
[24]https://onlinelibrary.wiley.com/doi/abs/10.1002/eco.1993
[25]https://royalsocietypublishing.org/doi/10.1098/rsfs.2016.0145
[26]https://www.journals.uchicago.edu/doi/abs/10.1086/708692?af=R
[27]https://royalsocietypublishing.org/doi/10.1098/rstb.2019.0361
[28]https://www.cell.com/trends/ecology-evolution/fulltext/S0169-5347(19)30043-6
[29]https://oxford.universitypressscholarship.com/view/10.1093/oso/9780199377176.001.0001/oso-9780199377176-chapter-10
[30]https://www.sciencedirect.com/science/article/pii/S0167198715300660
[31]https://www.sciencedirect.com/science/article/abs/pii/S0038071709003952
[32]https://www.sciencedirect.com/science/article/abs/pii/S0929139314002091?via%3Dihub
(參考文獻(xiàn)可上下滑動(dòng)查看)
復(fù)雜科學(xué)最新論文
集智斑圖頂刊論文速遞欄目上線以來,持續(xù)收錄來自Nature、Science等頂刊的最新論文,追蹤復(fù)雜系統(tǒng)、網(wǎng)絡(luò)科學(xué)、計(jì)算社會(huì)科學(xué)等領(lǐng)域的前沿進(jìn)展。現(xiàn)在正式推出訂閱功能,每周通過微信服務(wù)號(hào)「集智斑圖」推送論文信息。
原標(biāo)題:《演化工程師:生命與環(huán)境如何相互塑造》
本文為澎湃號(hào)作者或機(jī)構(gòu)在澎湃新聞上傳并發(fā)布,僅代表該作者或機(jī)構(gòu)觀點(diǎn),不代表澎湃新聞的觀點(diǎn)或立場(chǎng),澎湃新聞僅提供信息發(fā)布平臺(tái)。申請(qǐng)澎湃號(hào)請(qǐng)用電腦訪問http://renzheng.thepaper.cn。



- 報(bào)料熱線: 021-962866
- 報(bào)料郵箱: news@thepaper.cn
滬公網(wǎng)安備31010602000299號(hào)
互聯(lián)網(wǎng)新聞信息服務(wù)許可證:31120170006
增值電信業(yè)務(wù)經(jīng)營(yíng)許可證:滬B2-2017116
? 2014-2025 上海東方報(bào)業(yè)有限公司

